
From ancient times, people had noted the fossilized remains of animals that did not seem to resemble living species. Moreover, seashells could be found in the strangest places, even on the tops of the highest mountain ranges. The ancient Greeks were aware of these fossilized remains of creatures, and Heredotus (484-425 BC) suggested that they came about as a consequence of changes in the positions of the sea and land. These changes were even associated with considerable time periods, and Aristotle believed that they took place so slowly that they could not be observed today.
Many theories regarding fossils have been propagated, ranging from Lusi naturae “jokes of nature” to prehistoric animals buried by catastrophic events (adherents of this view included Rob- ert Hooke who discovered cells and Cuvier, the French compara- tive anatomist). Fossils were recognized as extinct species whose place has been filled by the creatures living today. Bible-believing scholars, who attributed the fossils to the destruction of animals during the Noachian flood described in Genesis, also accepted the catastrophic model. However, the gradual ascendancy of the idea of long ages, together with the numerous questions raised by the
Biblical account led many to doubt the validity of the Scriptures. Some of the questions that seemingly could not be accounted for were: How did all the animals get into the ark? Why is there a particular order in the fossil record? How did the animals get to the various continents from the ark? Why do the animals represented in the fossil record look so different from those present today?
These questions led to a search for naturalistic explana- tions for the fossil record and the origin of life in general. Before Darwin published his Origin of Species, Jean-Baptiste Lamarck (1744-1829) was one of the first of the new era of scientists to propose that the geological discontinuities in the stratigraphic record represented gradual changes in the environment and cli- mate to which species were exposed and through their effects on organisms these changes led to species being transformed. The geologists, Hutton and Lyell, expanded this concept and Charles Darwin added the biological arm, thus laying the foundation for modern concepts on the origin of fossils. Indeed, the fossil record is today considered to be the severest blow to all anti-evolutionary ideas.
Ironically, the scientific views on the question of origins have a tendency to go full circle. Whereas exponents of the theory of evolution rejected catastrophism, many scientists are today returning to catastrophism and even to the Biblical account of the flood to explain many of the features of the geological column and the fossil record. A major problem with this view, as scientists see it, is the universality of the Biblical flood with its destruction of all terrestrial life, which would put an end to any theory of natu- ralistic origins. However, it is not only the Bible that speaks about a worldwide flood, but virtually every society on every continent has the story of a worldwide flood in its folklore.1 Moreover, there is evidence that indicates that there was a universal total covering of the earth by water – compelling evidence that cannot readily be ignored. This includes:
- Massive fossil graveyards with evidence of plants and animals being washed into position.
- Huge sedimentary deposits (nearly three quarters of the earth’s exposed surface is covered with sedimentary rock deposits).
- The chalk deposits of the world are universal. Chalk is formed from the skeletons of marine unicellular protozoans and algae, and can only settle out of relatively shallow water. In deep oceans, the calcium carbonate shells dissolve on the way down to the ocean floor. The chalk deposits are thus an indication of worldwide coverage of a relatively shallow sea. Chalk deposits of the same age are found in many areas of North America, Australia, Europe, Asia, and Africa, and all of these deposits are resting on the same type of glauconitic sandstone.2 For these factors to be so universal, the same conditions must have existed universally.
- The vast coal and oil fields of the world are further evidence of a vast flood catastrophe. No process occurring today can even remotely approach the magnitude of the catastrophe necessary to account for such a vast scale of universal burial of plants and other organic material.
Order in the Fossil Record
The three eras of geological time (Paleozoic, Mesozoic, and Cenozoic) are characterized by different fossil assemblages (Refer to Figure 2.2). Before these eras, there was the Precambrian, which is not regarded as an era and contains fossils of blue-green algae and some fungi. The Paleozoic Era is known for its abundant marine life ranging from brachiopods and trilobites to sharks and bony fishes in the Devonian, as well as amphibians and reptiles in the Carboniferous. This era is also known for its extensive coal beds, consisting largely of extinct plants such as giant horsetails, ferns, some seedless plants, and club mosses. The Mesozoic era is divided into three periods, the Triassic, Jurassic, and Cretaceous, and is known for its dinosaurs and many other reptiles. At the close of this era, we have the extensive chalk deposits, which we have discussed earlier. It is not surprising then that the close of this era is associated with massive extinctions. In a catastrophic flood model, this era would be associated with the close of the flood period prior to the re-emergence of the continents from the waters which covered the earth. The plants of the Mesozoic era comprise cycads, conifers, and deciduous trees many of which still exist today. In the Cenozoic era, we find largely the fossils of mammals and birds, and Cenozoic plants are essentially similar to the species that exist today.
The question arises as to why there is this particular order in the fossil record which is interpreted as a progressive advance from simple to complex organisms? The type of fossil found in the various layers changes as one ascends the geological column, from invertebrates, fish, amphibians and reptiles, to the mammals and birds in the uppermost strata. This order in the fossil record is one of the prime evidences used by scientists to establish evolu- tion as a fact. However, the sequence is not from simple organ- isms to complex organisms (there is no such thing as a “simple” organism in biological terms), but rather from marine sessile to pelagic (free-swimming) to terrestrial life forms. The fact then is, that there is no such thing as a simple undifferentiated animal in the fossil record that may be cited as proof for the development of organisms from simple to complex. George Gaylord Simpson, the famous proponent of the evolution theory, already witnessed to this fact when he stated in his book The Meaning of Evolution:
It has been suggested that all animals are now spe- cialized and that the generalized forms on which
major evolutionary developments depend are ab- sent. In fact, all animals have always been more or less specialized and a really generalized living form is merely a myth or an abstraction.3
The earliest organisms in the fossil record were thus complex organisms and there is no evidence for the progressive advance required by the theory of evolution. Yes, there were many organisms such as trilobites and ammonites that existed in the past that do not exist today, but this does not make them primitive. They were just as complex as anything living today. In fact, the fossil record shows a past wealth of organisms which is staggering. Surprisingly, most of the organisms of the past were much larger and impressive than present-day animals even, where they belong to the same groups of animals. In fact, in this regard, the fossil record shows evidence for devolution rather than evolution.
There are many reasons beside progressive development which could be cited for the order in the sequence of fossils. The sequence from sessile to free-swimming to terrestrial is indicative of ecological zones being destroyed progressively as can be illustrated by a simple example. If a bulldozer rapidly covered a duck pond with soil, then the organisms in the pond would be buried in sequence. The bottom dwelling worms and snails would be at the bottom, the fish somewhat higher, and the ducks on top. The sequence represents the ecological zones in which the animals lived and definitely not the order in which they evolved. The same holds true for the fossil record. Indeed, we find assemblages of fossils e.g. radiolarians and foraminiferans in the same sequence in the fossil record as they occur in a present day ecological zonation.4 The ability to float or not would also produce zonation. Mammals and birds float due to bloating or trapped air in feathers and hair and are thus to be found in higher strata. Coal layers reflect this same phenomenon. The Paleozoic coal seams consist largely of non-floating plant species whereas
those found in higher strata consist of floating species. No model provides all the answers, but the flood model can provide a very satisfactory explanation for the series found in the fossil record.
Explosive Evolution
The bulk of the fossil record is found in a series of lay- ers commencing with the Cambrian. The Cambrian is associated with an explosive occurrence of various diverse fossil forms. The sudden and almost simultaneous appearance of fossils, from even different phyla, has been termed “the Cambrian explosion”. Pre- Cambrian fossils are largely limited to microorganisms, and where macrofossils occur, they are normally at contact zones with the Cambrian and difficult to ascribe to one or the other layer.
If evolution were true, one would expect a progressive advance from simple to complex in the fossil record, but amaz- ingly one finds that even the so called higher life forms such as the chordates appear right in the beginning. In an article entitled “The Big Bang of Animal Evolution”, Jeffrey S. Levinton, professor in ecology and evolution at the State University of New York states:
The Cambrian explosion was characterized by the sudden and roughly simultaneous appearance of many diverse animal forms almost 600 million years ago. No other period in the history of animal life can match this remarkable burst of evolutionary creativity.5
He goes on to say that evolutionary lawns rather than trees appear to be the norm. An evolutionary tree normally shows a common ancestor with branches leading to organisms that develop later in time. In a lawn, all the branches are parallel which means there is no so-called common ancestor. Levinton concludes:
Those stories point to a serious problem with all arguments about evolution that rely on taxonomic classification. Some of the fossils that suggest the existence of unique classes are very poor scraps from the geological table.
The fact that all the major phyla appear simultaneously in the fossil record is a strong argument for creation rather than evolution. Even Charles Darwin admitted this in the first edition of the Origin of Species. He writes regarding this issue:
The case at present must remain inexplicable and may be truly urged as a valid argument against the views here entertained.6
These facts have urged Neo-Darwinists such as Stephen
J. Gould to reconsider Darwin’s idea of gradualism (the slow development of one form out of another over time) and replaced it with the concept of evolution through punctuated equilibrium (periods of equilibrium followed by rapid (punctuated) changes). Whatever one wishes to term it, the fact remains that organisms appear suddenly which is in harmony with the creation account. Punctuated equilibrium has many hurdles to cross, particularly in the sphere of genetics, where it must be explained how so many useful mutations could come about so rapidly or be employed usefully in an integrated fashion without there being prior design. The idea that all major phyla could appear at once seems to be stretching it to say the least.
Current concepts require vast time periods, measured in millions of years, to accommodate changes from one life form to another. Standard geology, therefore, supposes vast time periods for each of the geological layers to account for this period of time. Historically, the time periods became longer as evidence
for the complexity of evolutionary change became greater. It is interesting that new evidence of the very rapid appearance of life on earth, and the absence of evidence for change over long periods of time, have forced scientists in the opposite direction, and they are just as willing to slash vast ages out of the geological column, as they were to insert them. 7
The problem is so vast that it almost seems as if scientists are becoming desperate to find a solution to the problem. They are even talking of “Evolving at Supersonic Speed”. A group of research- ers from M.I.T. and Harvard found it necessary to recalibrate the geological clock by chopping the time for the Cambrian in half and then cramming the evolutionary events into the first third, prompting Gould to state:
Fast is now a lot faster than we thought, and that’s extraordinarily interesting. 8
Gould states that full diversity was reached in the Cam- brian explosion, and this admittance is only one step away from special creation. Indeed creationists do just a little more chopping to the time scale. A recent discovery of even a vertebrate from the Cambrian shows beyond doubt that full diversity was reached in the Cambrian Explosion, and this further complicates the issue for naturalistic evolution over millions of years.9 If full diversity was there from the beginning, then that sounds like special creation.
Evolutionary Sequences
Gould states that:
The family trees that adorn our text books are based on inference, however reasonable, not the evidence of the fossils.10
This means that the evolutionary sequences are morpho- logical sequences and do not necessarily reflect the sequence in the palaeontological record. The sequences are logical according to the paradigm or mindset of the researcher, but they are not nec- essarily right. If a taxonomist were given the task of determining the evolutionary development of a series of dog skulls (given the scenario that he did not know how the various dog races came into existence and that the scientist had never seen living dogs), then he or she would logically group them from small to large and also group the flat-nosed ones on one branch of the evolutionary tree and the long-nosed ones on the opposite branch with a com- mon ancestor somewhere at the base of the tree. This would be perfectly logical, but it would not be right, since dogs all belong to one species and the variation between them is as a result of the built-in variation in the original gene pool. The dog was not bred from the smallest to the largest, but these reflect the extremes of the genetic variation within the gene pool. The dogs were in fact bred from the center outwards with wild canids as a starting point. The palaeontologist suffers similar constraints as our hypothetical researcher. He or she is also confronted with the bones of animals often from the same strata and asked to arrange them without hav- ing seen the living specimens and not knowing their origin.
Given the nature of the fossil record, many species would have been contemporaneous which means that they lived at the same time. Scientists accept that a species may evolve and then continue to exist unchanged whilst other species may have evolved out of this species (or similar species), the coelacanth being a case in point. However, the other possibility is that the coelacanth and other so-called living fossils such as Solenodon, a shrew-like mammal that disappeared from the fossil record some apparent thirty five million years ago, the tuatara lizard from New Zealand of which there is not a trace in the fossil record since the Mesozoic, and Lingula, the marine creature that was to have become extinct some five hundred million years ago are just what they represent, creatures that were there from the beginning and still exist today.
In fact, unchanged (except for size) scores of insects and inver- tebrates in general and hosts of plants have unchanged records from their first appearance in the fossil record to the present. The perfect preservation of many soft bodied animals such as jellyfish and the superb preservation of insects and crustaceans show that even down to the finest detail the fossil forms resemble the living creatures of today.
The fact that they appear and remain unchanged could thus be an indication of design, but this is generally too simple a solution for those seeking a naturalistic answer to origins and so, given the contemporaneous nature of many species, scientists are compelled to piece together the evolutionary puzzle from what is available to them. The fossils are then arranged in sequences consistent with their morphology and the paradigm of the researcher. Using modern bio- chemical techniques, it is now possible to test these fossil sequences using DNA comparisons between species. Interestingly, this often confuses the issue even more, because vastly different creatures on the morphological level often turn out more related than morphologi- cally similar creatures. Also the sequence of development is brought into question by such determinations. For example, cartilaginous fishes, such as sharks, are considered to be more primitive than the bony fishes, but comparison of the complete mitochondrial DNA sequence of the spiny dogfish indicates that it falls between the teleosts and non-teleost bony fishes on the phylogenetic tree. This implies that cartilaginous fish have lost the swim bladder and the bony skeleton and may thus not be as primitive as supposed, but rather regressed. 11
A study of the major living groups of fishes and their sup- posed ancestry shows that their ancestral lines do in fact not link up to form a tree, but can be traced back in parallel lines (evolutionary lawns), often unchanged except for size, without any links to other life forms. Intermediaries are also missing and links are thus based on speculation, not the evidence of the fossils The same holds true for amphibians, reptiles, birds, and mammals.
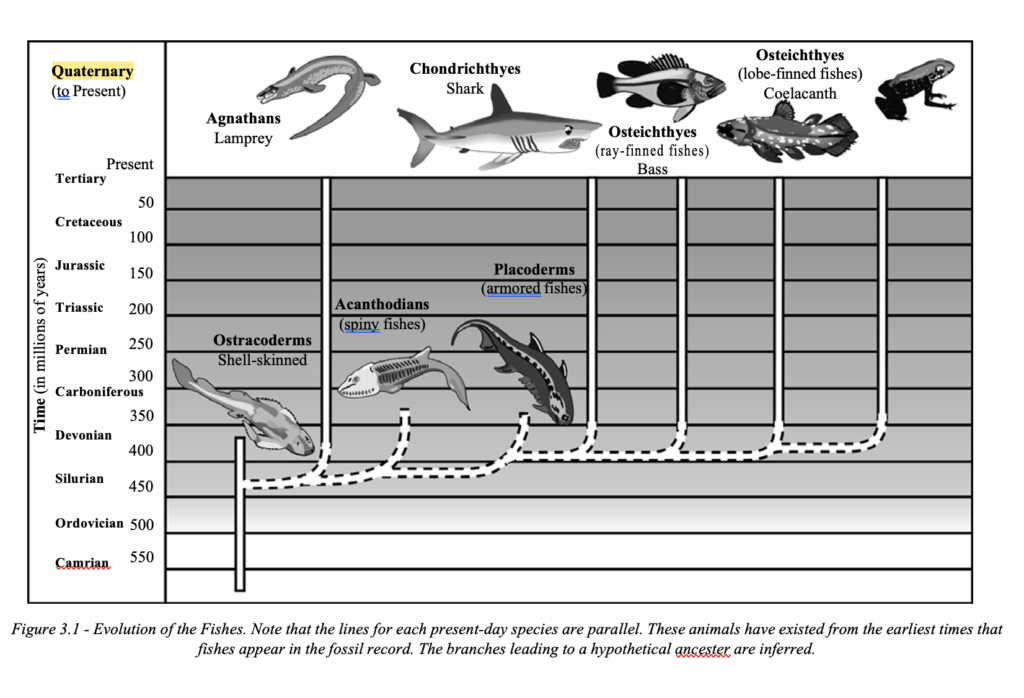
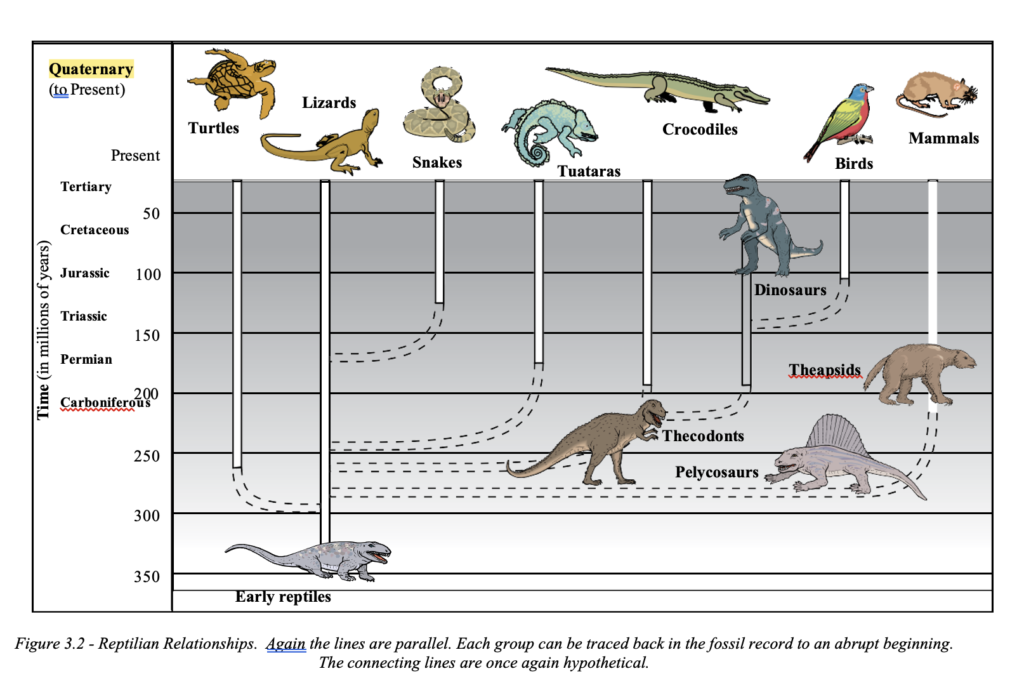
Their lineages are traced in parallel lines in the fossil record and hypothetical evolu- tionary trees are thus based on inference (See Figures 3.1 and 3.2).
Intermediaries
Intermediaries in these supposed evolutionary lines are absent, although creatures such as Archaeopteryx and the mammal- like reptiles are cited as evidence for links between reptiles and birds and reptiles and mammals. The mammal-like reptiles suffer from the same problem as the other sequences, since again they are based on morphological sequences as in the case of our dog scenario. Logical sequences can be presented, but this does not necessarily make them right. Arranging these creatures according to skull morphology suffers the same constraints as arranging the dogs on the hypothetical tree according to their skull morphology. In a sense, there is thus an element of circular reasoning, since one is using a morphological sequence to back a second morphological sequence that is also based on inference. As the theory of evolution requires links, however, scientists are obliged to fill in these gaps with hypothetical intermediaries, and these morphological sequences are used for just this purpose when in fact they can at best remain hypothetical missing links.
Archaeopteryx, for example, was found two years after Darwin published his Origin of Species. This fossil has reptilian and bird features, which are not unique to either group, but there is a measure of controversy regarding the fossil, since some consider it an artifact or even a hoax.12 Generally, the fossil record does thus not provide the expected intermediaries that are required to fill the gaps in the record. As the erstwhile Professors of Zoology at Harvard University, A.S. Romer and George Gaylord Simpson, admit:
‘Links’ are missing just where we most fervently desire them, and it is all too probable that many ‘links’ will continue to be missing.41
It remains true, as every palaeontologist knows, that most new species, genera and families – and that nearly all new categories above the level of families – appear in the record suddenly, and are not led up to by known gradual, completely continuous transitional sequences. 42
Even Dr. Colin Patterson, senior palaeontologist at the British Museum of Natural History admitted in a letter written in 1979, in answer to a question regarding the absence of intermediary fossils in his published materials:
I fully agree with your comments on the lack of direct illustrations of evolutionary transitions in my book. If I knew of any fossil – or living, I certainly would have included them. You suggest that an artist should be asked to visualize such transformations… but where would he get the information from? I could not honestly provide it, and if I were to leave it to artistic license, would that not mislead the reader?
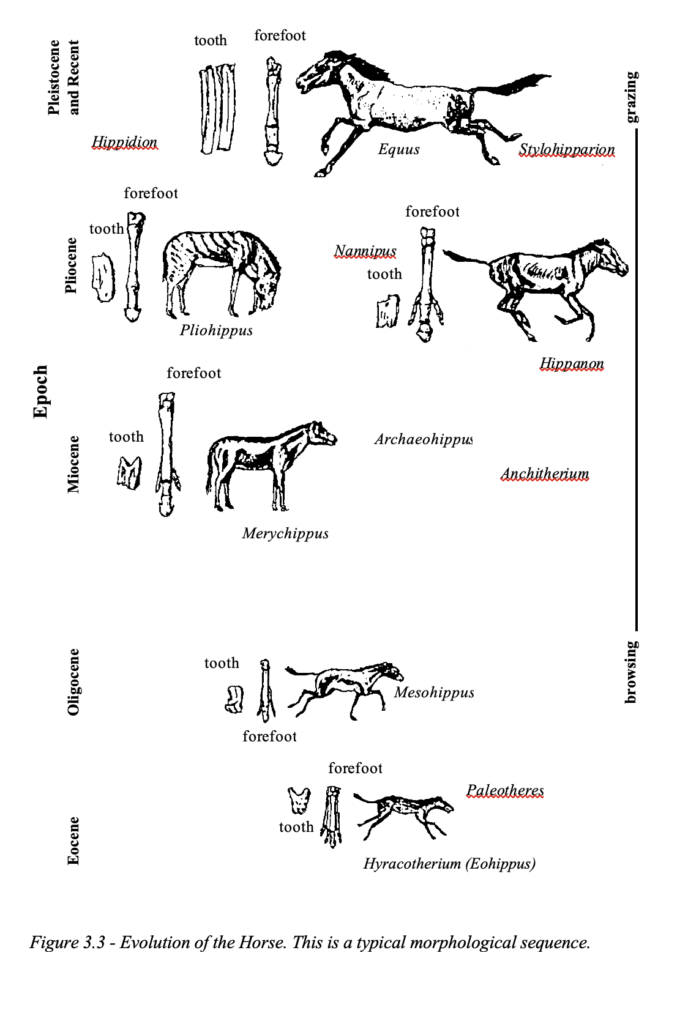
Evolution of the Horse
Probably the most famous example cited in support of the gradual change of species over time is the evolution of the horse. This exhibit is placed in virtually all textbooks dealing with evolution and it receives pride of place in the museums of the world (Figure 3.3). Once again, we are dealing with a morphological sequence, worked out by O.C. Marsh13, of contemporaneous creatures ar- ranged from small to large. There is no evidence linking them in a continuous chain of transitional forms and controversy exists as to finer transitions such as the reduction of the number of toes as we move from Eohippus (which was previously called Hyracotherium and resembles the rock hyrax) to the various intermediaries to Equus, the modern horse. Eohippus was discovered in 1841 in clay around London. It did not even resemble a horse, and it had sharp as well as multiple back teeth as in hoofed animals. Of course, be- ing a morphological sequence these different fossils are found in the same strata, sometimes right next to each other. Marsh already knew that some modern horses have additional toes, which makes them similar to the extinct Protohippus.14 Moreover, the various species used in the horse lineage have variations in the number of ribs (they vary from 15 to 19 and Equus has 18) and lumbar vertebrae (vary from 6 to 8), which would require genetic reversals contrary to the evolutionary model which states that evolution cannot be reversed. Variation in horse size is not an indication of evolution, since we have large variations in the size of living breeds today. The largest horse living today is the Clydesdale and the smallest is the Flabella which is only 43 centimeters (17 inches) tall and both, of course, belong to the same species.
In my own experience when lecturing on the question of evolution and creation, I have been amazed that palaeontologists are fully aware of these details, and in one lecture given at a promi- nent South African University, one of the leading palaeontologists frankly admitted that: “No one believes that anymore.” Why is it then that the exhibits remain in the museums when ‘no one be- lieves that anymore’? The speculative evolution of the horse is thus acknowledged by the scientists’ themselves15, and even Simpson admitted that:
The most famous of all equid (horse) trends, ‘gradual reduction of the side toes’, is flatly fictitious.16
Marine Mammals
Marine mammals are an enigma, since they show such perfect adaptations to a marine existence in both their anatomy and their physiology. Marine mammals are supposedly derived from eutherian land mammals, which secondarily reinvaded the sea and evolved to their present state in the short period (even in evolutionary terms) available since the dawn of mammals. In order to achieve the remarkable capacity to swim and dive as they do, some fascinating physiological changes had to take place. In the case of whales and porpoises, the hind limbs had to disappear and the posterior portions of the animals had to be transformed into a fin together with the associated muscular design capable of producing the forces necessary for the type of explosive swim- ming for which these creatures are known. Evidence for such an evolutionary development is lacking in the fossil record, although such profound changes should surely have left their mark in the stratigraphic record.
There have been attempts to explain the origin of whales from the fossil record, but most fossils show that they looked just like present-day whales. Variations in the position of the respira- tory opening are used to show evolutionary progression, but these progressions are again based on morphological sequences and not on any sequence of the fossils and they therefore at best reflect normal variation. Evolutionists, however, tend to use three fossil creatures as evidence for whale evolution, these being Pakicetus, Ambulocetus, and Basilosaurus. Of these, Pakicetus has been reconstructed as a walking whale from a few fragments of jaw and skull. The creature was a land mammal, it was found buried with land mammals and there is no evidence to link it with whales except in the mindset of the scientists. Ambulocetus is a creature known from some skull and limb fragments, which is again used as an intermediary, but since true whales already existed together with this creature, we are again dealing with morphological sequences, not the evidence of the fossils. Moreover, no hip was found with this creature, making the entire conjecture as to its lineage even more speculative. Basilosaurus is an odd serpentine creature, which has little to do with whale evolution and as some scientists freely admit, they could not possibly have been ancestral to modern whales.17
Seals, sea lions and walruses present just as great an evo- lutionary enigma since they also make a sudden appearance in the fossil record. All these animals are grouped under the name pine- peds (fin-footed) and are divided into three groups, the Phocidae, Otariidae and Odobenidae. The Phocidae are the so-called ‘true seals’, the Otariidae are the ‘eared seals’, and the Odobenidae are the walruses. Fossil seals appear suddenly in the fossil record and look very much like those living today.18 In fact, the similarities are so great, that monk seals are considered by some to be living fossils because of their unchanged status.19 The common features shared by the three groups has led to some interesting debates as to the ancestry of these animals, with some scientists suggesting different ancestors ranging from bear-like to otter-like creatures, but there is no evidence for such a convergent evolution and there is no fossil evidence to back it up.20, 21
Catastrophism and the Fossil Record
Evidence for catastrophism is very widespread in the fos- sil record, and the mere fact that most fossils are embedded in deposits laid down by water thus having had to have been buried rapidly, indicates catastrophic formation. Geologists have long recognized that:
If covered by moist sediment, weathering is prevented. For these reasons quick burial is perhaps the most important condition favoring fossilization.…. Water borne sediments are so much widely distributed than all other agents of burial that they include the great majority of fossils. 22
Some fossils show immaculate preservation of detail and these creatures must have been buried instantly to prevent decay. The state of preservation can thus be an indication of how long the animals were exposed to the elements before they were bur- ied, and in a flood scenario, it is envisaged that some creatures would have remained uncovered longer than others. In fossil fish for example, we find perfectly preserved specimens, specimens without heads, with and without scales, and sometimes just the bones or pieces of specimens. During putrefaction, the scales and the heads of fish drop off quite rapidly, and so it is evident that some were buried instantly and some floated for some time before burial. There are four major types of fossils depending on the method of preservation. These are molds and casts, comprising footprints and molds that have been filled in with rock-forming material, petrified fossils or fossils turned to stone by replacement of the tissue with the elements of the surrounding strata, carbon- ized fossils, such as coal, and, lastly, unchanged fossils such as animals trapped in amber or the preserved parts of animals trapped in tar pits. Seashells and tooth and bone fragments also belong to the latter group.
Fossil Footprints
In the Grand Canyon series, as in other areas of the world, there are parts of the stratigraphic record which scientists ascribe to periods of aridity and are thus supposed to be desert deposits. The Coconino Sandstone is one such deposit in the Grand Canyon series and it consists of fine-grained quartz sand and is up to 1000 feet thick in some areas. It is also crossbedded which is common of either desert or underwater dunes. The sandstone layer also contains tracks made by vertebrates and invertebrates and most of them tend to have been made whilst the animals were moving uphill in terms of the crossbeded layers. This is, in fact, one of the features of most of the trackways and warrants an explanation. Moreover, a desert period in what creationists would consider to be flood deposits would also be problematic. The issue of desert deposition has received some new attention from scientists, and on closer examination, it has been found that underwater deposi- tion is a more likely model to account for their features because they are more in line with depositions that have been studied in shallow marine sediments than those that could be formed under desert conditions. These authors state:23, 24, 25
Since 1903, most of the Navajo sands were assumed to represent ancient wind dunes formed on a vast Sahara-like desert; this became a ruling hypothesis…. The Navajo problem originated years ago when geologists could conceive of large amplitude cross stratification as originating only in wind-formed dunes; no other modern processes that could form it had been studied. This highlights the major shortcomings of reasoning by analogy, namely the limitation at a given time of known possible analogues. Today, knowledge of modern shallow marine sedimentation has broadened the spectrum of counterparts and analogues. Insight gained into remarkably large underwater dunes
found in very shallow shelf areas provides as attractive a comparison for much of the Navajo sands as for lower Paleozoic quartz sandstones.23
Inasmuch as geologists are forced to interpret ancient sediments chiefly by analogies with modern phenomena, interpretations are severely biased if all possible modern analogues are not known; such was the case when the Navajo was first studied.24
Studies of the actual footprints in the Coconino Sandstone and comparisons with footprints made by animals in laboratory conditions on various substrates showed that the tracks in the sand- stone must have been made in wet sand consistent with underwater formation and that the animals made similar tracks when walking upgrade.25 There are two possibilities here. Either the animals do not leave adequate tracks when going downhill, or they were go- ing uphill for a reason. Laboratory experiments show that animals leave equally good tracks when going downhill as when going uphill, and so the only other logical conclusion is that the animals were going uphill because they were perhaps trying to escape rising water levels. This is consistent with the evidence, and also fits the catastrophic flood model.
Collating data of fossil footprints (from about 800 pub- lished papers) and the actual presence of the body fossils of the creatures that could have made these tracks in the same strata has also yielded some surprising results. One would expect that strata containing footprints would also contain the fossils of the animals that could have made them, but this is not always the case.26 The distribution of bird and mammal footprints correlate well with the distribution of body fossils, but amphibian and reptile footprints correlate poorly with body fossils. Amphibian footprints are rare after the early Permian and reptile footprints, with the exception of dinosaur footprints, are most abundant in late Triassic and early
The Genesis Conflict
Jurassic rocks. The only Cretaceous reptile footprints identified in the literature are about a dozen types of dinosaur footprints. Now in contrast, the amphibian and reptile body fossils are the most abundant in the Cretaceous and the Tertiary where corresponding footprints are rare or nonexistent. This does not seem to make sense if the geological column represents strata accumulated over millions of years, since one would expect the actual creatures to abound in the same time period (strata) where their footprints are recorded. However, if the strata were deposited catastrophically and do not represent millions of years of time, then a logical explanation can be found for this anomaly.
Mammals and birds would not readily make underwater footprints since they tend to float in deep water, and the larger mammals and the birds could have sought high ground during the initial flood events. During the early flood events, large numbers of amphibians and reptiles would have been moving about and thus producing footprints, which would have been preserved by being covered by rapid deposition mud flows. Later in the flood very few live reptiles and amphibians would produce footprints, except for the larger creatures such as the dinosaurs. During the Cretaceous, when the only footprints preserved were those of a few dinosaurs, there would have been many amphibian and reptile bodies that were being buried by the catastrophic formation of these strata, and this would account for the abundance of body fossils of these creatures that are found in these layers. Footprints of ‘bird’ tracks have been found in deposits in Nova Scotia, but since these are carboniferous deposits, they are presumed to have been made by other creatures, since birds should not have been around yet. If the same prints had been found higher in the column, then they would have been classified as bird prints. Similar tracks have also been found in Permian deposits.26
Generally speaking then, the footprints in stone support the catastrophic model better than the conventional model of long ages.
Dinosaurs in Mud
Since fossilization requires very specific, complex condi- tions, the vastness of the fossil record does not support the idea of uniformitarianism. Indeed when one considers the giant dinosaur fossils, then the question arises “What buried these creatures rapidly?” Dinosaurs are sometimes found in relatively large numbers and mostly they are washed into position with many of them showing distinct stream orientation. Scientists explain this by stating that these creatures lived in the flood plains and were periodically overcome. However, these fossil beds often stretch over thousands of square kilometers, which would not be consistent with such a scenario. Moreover, some dinosaurs have been found imbedded in marine deposits, suggesting that they were washed out to sea. Recently, a specimen of Scelidos- aurus with excellent preservation of even skin tissue was found in Great Britain in marine deposits together with algal spores and bivalves.27
Dinosaurs were of course reptiles (giant lizards), and their lineage cannot be determined from the fossil record. However, they have received such media attention because of their size and presumed ferociousness that they have captured the attention of young and old. Far from proving evolution, they are in fact one of the stumbling blocks, since animals of such remarkable diversity in size and form must have had a recognizable lineage. The heavi- est dinosaur on record is the estimated 100-ton Argentinosaurus, but it was not the largest, as some sauropod specimens have been found that could have been 50 meters long and standing 14 meters high. Large numbers of dinosaur eggs, apparently from these large creatures, have also been found in Argentina, and these are being interpreted as dinosaur nurseries, but again these well preserved eggs were buried rapidly in silt from a flood.28 Chiappe, one of the team leaders and principle author of the paper published in the journal Nature had this to say regarding the eggs:
The Genesis Conflict
Scientists found so many embryonic remains that it appears catastrophe struck the nesting ground, keeping many eggs from hatching … Floods may have penetrated the porous shells and drowned the embryos. 29
There are some that say that dinosaurs are not extinct, but live on in the birds of today. The evolution of birds is traced to dinosaurs, but there are major problems with this supposition. The feathered dinosaur Archaeopteryx resembles a small coelurosau- rian dinosaur Compsognathus and the only distinction linking it to birds is its feathers, which have been disputed in the literature as previously noted. Other than the disputed feather link, there are other problems, which negate dinosaur lineage for birds. Birds lack the embryonic thumb that dinosaurs had, suggesting that it is “almost impossible” for them to be related.30 Moreover, it is impossible to envisage how the simple sac-like lung of dinosaurs could have evolved into the highly complex avian lung with its through-flow system and countercurrent oxygen exchange. A team led by John Ruben, from Oregon State University, analysed outlines of Sinosauropteryx (a so-called feathered dinosaur) and concluded that its “bellowlike lungs could not have evolved into the high-performance lungs of modern birds”. 31
There is no denying that floods and fossils seem insepa- rable, but an erstwhile world destroyed by a flood would destroy the very foundations of the evolutionary theory, which sees fossils as evidence of phylogeny. However, more and more evidence is coming to light which threatens the very foundations of uniformi- tarianism. One of the finest modern-day detective stories regarding catastrophism in the fossil record is to be found in the story of the petrified forests in Yellowstone National Park and other petrified forests around the world.
Petrified, Fossilized Trees and their Age Implications
The general distribution and vertical stratification of the petrified trees in the Yellowstone National Park are presumed to represent a series of up to 40 successive forests, one on top of the other, whose combined age was estimated as being well in excess of time-restraints imposed by a flood model. It was believed that each successive forest was covered by volcanic ash in sequential eruptions, only to be replaced in the course of time by new forests. It was argued that this evidence could not support a young age for the current topography of these forests. Dr. Harold Coffin carried out a detailed investigation of these petrified forests and discovered that in spite of the long age paradigms accepted in the scientific community, these forests actually support the catastrophic model.32 As many of the trees are standing upright in an apparent position of growth, it was accepted that they were the remains of an actual forest. Moreover, the strata in which each forest layer is situated are uniformly flat and totally unlike any modern forest which, if it were covered by volcanic ash, would show trees grow- ing on slopes and other uneven topography. Closer examination reveals that the petrified trees have no bark, no side branches, and the rootstocks are ripped off. This is inconsistent with trees being covered with ash while in a position of growth, since the roots would be undisturbed if the tree was still in position of growth.
Moreover, covering the trees with ash would certainly leave the branches at least in the vicinity of the trees that were covered.
There are not only upright trees in these strata, but many horizontal trees are also found. In places, the vertical separation is actually very small – the new layer lying just above the stumps of the older layer. Close examination of the strata reveals typical evi- dence of turbidite action and sorting of layers, which tend to show reverse grading with the coarser material on top. This is consistent with material which has been deposited by water-induced slides and slumps, and does not support deposition of dry volcanic ash.
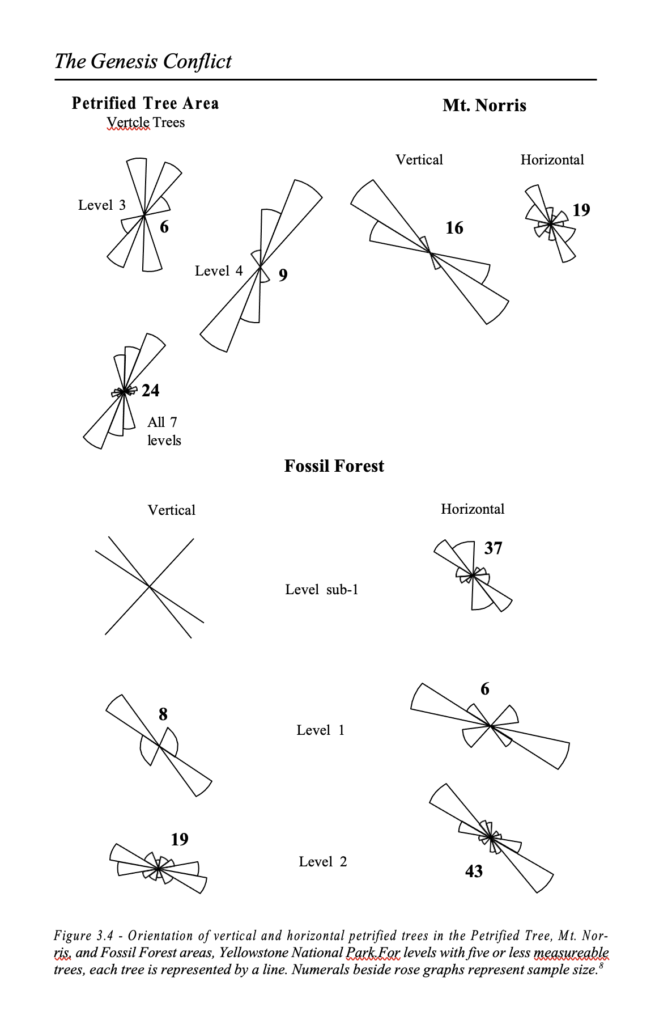
3 – The Fossil Record
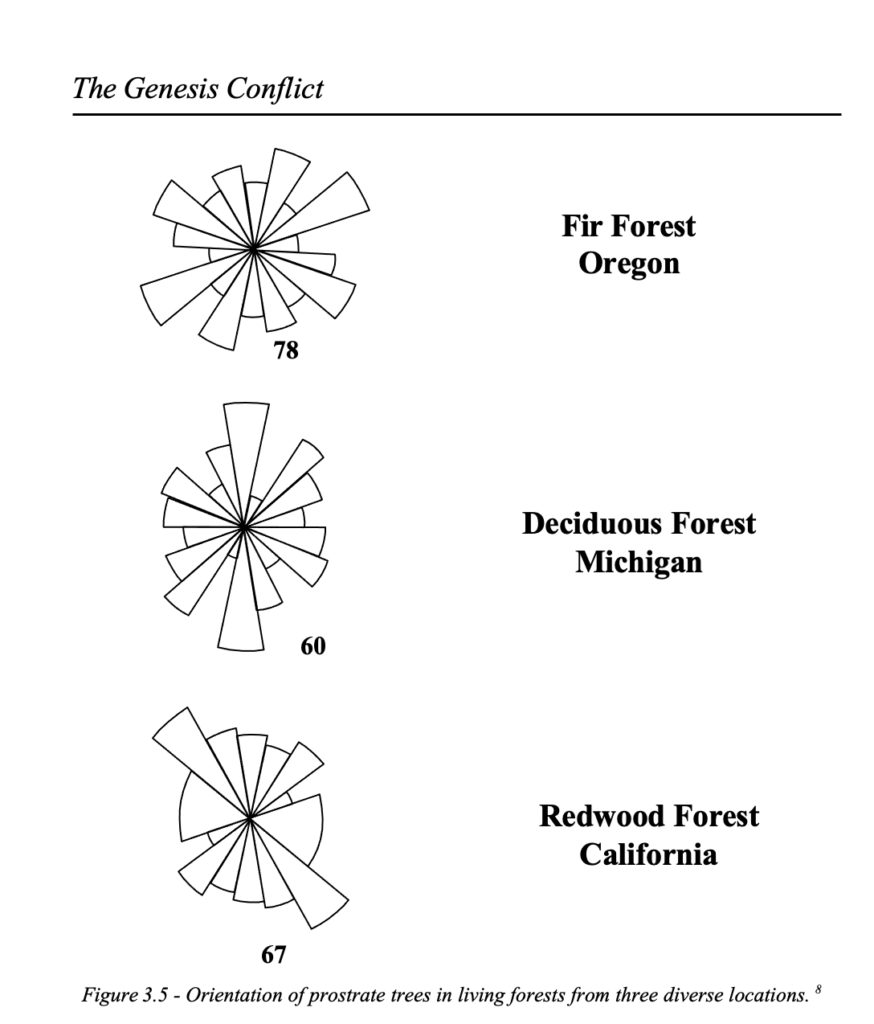
The organic layers, which previously were considered to represent the compacted forest floors, are also water sorted, which belies a mere forest situation. Analyses of tree orientation show that both horizontal and vertical trees are orientated in distinct directions. By comparison, the orientation of fallen trees in standing forests in Oregon, deciduous forests in Michigan and redwood forests in California show a lack of orientation (figures 3.4 and 3.5).
A solution to these anomalies came when, in 1980, Mount St. Helens erupted. The March 30 eruption melted the glacial ice, precipitating a flood on the south side of the mountain. Along with cold volcanic ash, the rushing water carried a large number of trees down the side of the mountain. These trees of varying sizes were stripped of their side branches, bark, and roots. The logs were buried in the volcanic ash with a predominant stream orientation. This is similar to the orientation of the petrified trees on the slopes of Mount Horniday in Yellowstone National Park. On May 18, Mount St. Helens erupted again, with an ac- companying earthquake. Tremendous pressure within the mountain was released after a rockslide, and the top 400 meters of the moun- tain were blown off in a catastrophic explosion. A force equivalent to 500 Hiroshima atomic bombs was unleashed. The destruction of the forest was total, with the trees literally blasted out of the ground. Debris falling in the lakes surrounding the mountain, caused tidal waves which washed uprooted trees into newly formed and existing lakes. In Spirit Lake, a study was done, and it was found that those logs that had root stumps rapidly righted themselves assuming a vertical position. A sonar scan of the bottom of the lake revealed 19,500 upright trees on the bottom of the lake.33 If one applies this scenario to the situation prevailing in Yellowstone National Park, one can readily account for the existing situation on the
basis of the catastrophic model.
More than one eruptive cycle would cause numerous turbidity currents and account for the numerous layers super- positioned over each other. Moreover, studies on the chemical composition of the volcanic deposits show that they were from eruptive events occurring simultaneously over a short period of time.
This can be ascertained because the relationship between chemical components in volcanic ejecta is constant only for single eruptive cycles. Studies on lava flows in Hawaii show that eruptive events separated by more than three months can be distinguished on the basis of the magma composition. The time implications for the formation of the petrified forests are thus consistent with a short chronology. Further evidence for catastrophism can be found in the fossil trees that are embedded in more than one geological layer simultaneously – a situation which is impossible if uniformitarian principles are applied.
How long would it take for wood to petrify? It has always been a tenet of evolutionary thinking that stone formation and processes such as petrification must have taken place over mil- lions of years. However, as we have seen in the previous chap- ter, these processes can take place rapidly. Instant petrification has been achieved artificially and under natural circumstances. By impregnating wood with solutions high in minerals such as silicon and aluminium, instant petrification has been achieved and even patented.34,
Writing in the Australian Lapidary Magazine, Piggot recounts a story of natural rapid petrification of a piece of wood from a tree that had been chopped down 70 years ago, with axe marks still on it, that had been buried and dug up again petrified. The story also reports on petrified fence posts with drill holes and wires still attached.35 Given the right circumstance, it would thus not take long for petrification to take place. Moreover, floodwaters together with volcanic ash would provide the perfect mineral soup for the processes to take place.
Floodwaters Covered the Earth
A catastrophe of the awesome magnitude proposed by the Biblical flood model would have totally restructured the post-flood world. According to Scripture, the whole world was submerged un- der water, and the restructuring of the earth to produce the present topography must therefore be a post-flood phenomenon. Evidence for total submersion of the continents is widespread on earth. Water deposition is a feature of the geological column, but one layer in particular, the Cretaceous layer, points to a transition between the pre- and post- catastrophic events postulated in this model.
The Cretaceous layer comprises of chalk deposits consisting largely of calcium carbonate derived from vast deposits of coccolith (algal) shells and other microorganisms with calcium carbonate skeletons. In view of its universal distribution, the Cretaceous layer
is evidence of a worldwide shallow sea covering the continents. The calcium carbonate skeletons of certain algae and Foraminifera would only settle out in large quantities if the seas were shallow and conditions favored algal blooms. Such disturbed ecological conditions would have prevailed in the immediate post-flood era.
The Cretaceous layer varies in thickness, a condition that could have been brought about by currents, or by differences in the time that the various areas were submerged under water. In the area of the white cliffs of Dover, the deposits are substantial, possibly indicating that these areas were submerged for a long period. This type of deposition does not occur today, as the calcium carbonate skeletons would dissolve in the deep oceanic waters presently existing. Although no present day scenario can parallel that of the deluge model, there are, however, some events occurring today which can shed some light on what might have happened in the past. The present disturbed ecology has resulted in some extraor- dinary algal blooms in waters rich in inorganic salts, derived from agricultural endeavors or other chemical industries. One such area is the Mediterranean, where masses of effluent and chemicals provide environments conducive to massive algal blooms.
The post-flood waters would have been rich in minerals and decaying organic materials, and in such circumstances, the algal blooms which produced the chalk layers could have been deposited in a very short time. Continental uplift would then have resulted in the drainage of water, recycling of sedimentary deposits and sub- sequent burial of the chalk layer, plant debris, and decayed animal remains. Further evidence supporting this model can be found in the Tertiary deposits which are packed with fossils creating fossil graveyards with numerous species dumped together. Strata with pieces of broken mammalian bones are also not uncommon in Cenozoic deposits, a condition that is difficult to explain using the standard evolutionary paradigm. Furthermore, stream-orientation of fossils is evident in the Tertiary which shows that deposition of these fossils was not only produced by water, but that the deposition was also catastrophic.
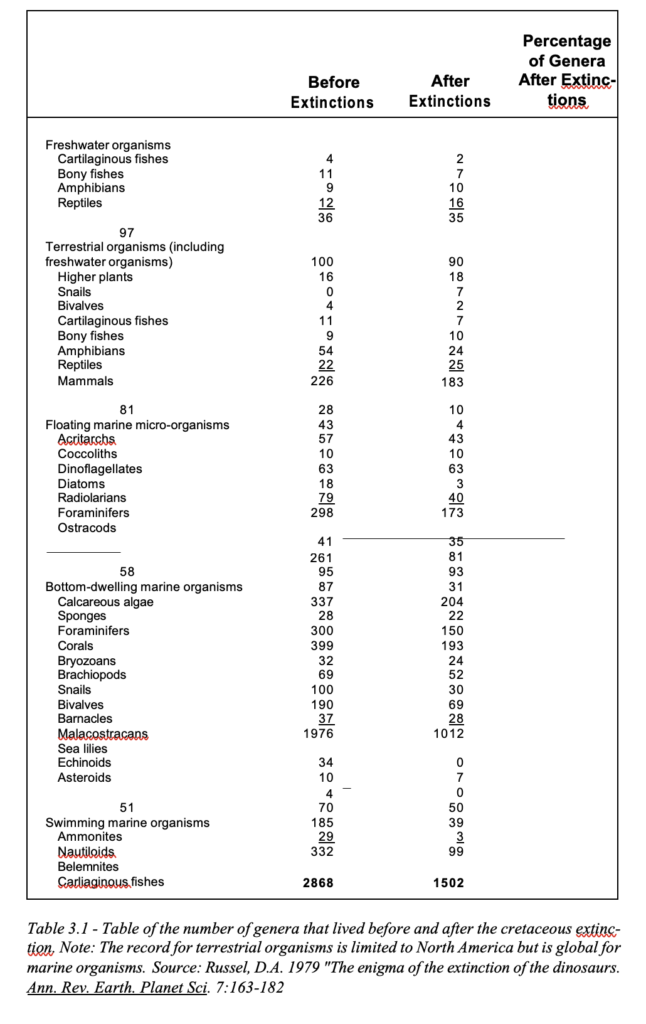
A catastrophe of this magnitude must surely have left its mark in the stratigraphic record. Indeed, the later cretaceous is as- sociated with huge scale extinction of numerous species including the dinosaurs. An analysis of the genera that survived this great extinc- tion at the end of the cretaceous shows that, besides the complete extinction of the dinosaurs, more than 50% of marine organisms also died in the destruction. In fact, the post cretaceous world is a shadow of what it was prior to this time period. The ammonites and belemnites suffered complete destruction, of all the swimming reptiles, only three survived. In fact, only 30% of all swimming marine organisms survived, whereas the survival rate of the fresh- water organisms (97% survival) was much higher (see Table 3.1)
Marine organisms are adapted to stable conditions, and a large-scale upheaval of the marine environment can be expected to lead to large-scale destruction. Numerous fossil beds of redis- tributed corals and mollusks account for massive destruction of the once stable marine environment. It is therefore not surprising that only approximately half of the bottom dwelling marine organisms survived this event. In the light of this destruction and the large- scale KT extinctions, it is not surprising that scientists have spent so much time debating the reasons for this phenomenon. Among the hypotheses suggested to account for extinction on such a mas- sive scale are intense volcanic activity, epidemics of disease, large scale greenhouse effects with a rise in CO2 levels leading to the death of dinosaur embryos, changes in plant composition, change in ocean salinity, high ultraviolet radiation, dust clouds caused by collisions with comets or asteroids, and ionizing radiation from supernova explosions.
Most of these theories concentrate on the dinosaurs, but fail to explain the large-scale destruction of all the other life forms. Surprisingly, a worldwide destruction by water comprising large scale upheaval of the ocean floor and submergence of the continents is totally absent from all the scientific conjectures regarding this era of extinction when all the evidence points precisely to such an event. The chalk bed deposits of the cretaceous period (‘Creta’is the Latin for ‘chalk’) are proof that everything was under water.
Sometimes it is difficult to see the forest for all the trees.
Surviving the Catastrophe
If one were to seek a reason for the extinction of many of the great creatures that once roamed the earth, then one might conjecture that the post-catastrophic world is not conducive to their survival. Firstly, there is evidence for a massive increase in the salinity of the oceans. As a comparative physiologist, I have always been fascinated by the fact that marine fishes (both the cartilaginous fishes as well as the bony fishes) are anatomically and physiologically adapted to a fresh water environment. Their internal salt concentration is approxi- mately one third of that of seawater and their kidneys are adapted for the elimination of water (they retain glomeruli which would normally form ultrafiltrates, but in the case of marine fish they have become functionally aglomerular by permanent constriction of the afferent glomerular artery), although this function is not required in seawater. In fact, their low salt concentration causes them to lose water by osmosis so that they cannot afford to lose water via the kidneys. The cartilaginous fishes (sharks and rays) solve this problem by retaining urea (a toxin) to raise their osmolarity to a level higher than that of seawater so that they can gain water by osmosis, whereas bony fishes desalinate the seawater by making use of salt pumps in their gills. Obviously these organisms were adapted to much lower salinities in the past and only survive because of their ability to osmoregulate under these circumstances. The retention of toxin by a cartilaginous fish is an indication of an emergency solution to which they eventually adjusted. Only organisms that could either conform to the new conditions or regulate their salt content could survive and the more sensitive are now extinct.
One might question the possibility that marine and fresh water organisms could have survived if the waters of the earth were all dumped together during a global flood. The separation between
fresh and salt water would then only be re-established once the continents had risen above the water level. Surprisingly, however, large bodies of fresh and saline waters, or even large bodies of fresh waters from different sources can coexist side by side without much mingling along the contact zones. This is seen in the great Amazon River, where two bodies of water run side by side for kilometers on end with limited mingling at the contact zone. During the flood stage, organisms adapted only to fresh water would thus have been able to survive in large bodies of fresh water that would have remained relatively distinct. Moreover, the marine life is enhanced where great bodies of fresh water come into contact with the ocean, and many species can only spawn in fresh or brackish water thus indicating that these were the conditions to which they were earlier accustomed. Of course, one would expect large-scale destruction of aquatic life in areas where the water composition was rapidly and radically changed due to hypersalination from underground aqueducts and through catastrophic mingling of the water masses, and this is indeed what we do find. The fact that so many marine organisms use rivers and estuaries as their spawning grounds indicate that the best survival salinities for these creatures must exist under these low saline condi- tions and that is why they will even migrate long distances to spawn in such areas.
Regarding the land organisms, it is enlightening that there are only two categories of organisms, in terms of their ability to cope with the thermal environment, in existence today. Terrestrial animals are either endothermic or ectothermic. Endothermic animals (largely mammals and birds) control their body temperatures by increasing their metabolic rates when environmental temperatures drop, whereas ectothermic animals control their body temperatures by selectively utilizing external sources such as solar radiation and environmental heat fluxes. In the absence of solar radiation, the body temperatures of these animals are the same as that of their environment. We also speak of these categories as warm-blooded and cold-blooded animals. In a world with climatic extremes, one would have to belong to either one of these two categories to survive.
There is evidence that the great reptiles of the past were probably neither endotherms nor ectotherms, but somewhere in between. The same probably holds true for many of the now ex- tinct giant amphibians and mammal-like reptiles. Studies of bone to marrow ratios show that the dinosaurs and other creatures were thus in this intermediary condition and would thus require stable environmental conditions. The plant life of the lower stratigraphic record shows that the earth probably had a relatively warm climate prior to the catastrophe, and the post-catastrophic climate was not suitable for the survival of the paleoforms. Moreover, the large- scale reduction in vegetation associated with the destruction means that many food sources were no longer available and precludes sur- vival of many of these animals. The paleontological record shows that far greater varieties of plants and animals existed in the past than are living today and that at both the plant and animal level, we are thus deprived of numerous species that must have once graced the planet. One could also expect that only non-specialist feeders could survive the destruction of their preferred food source, so that many of the great creatures of the past are no longer with us because the planet does no longer provide their niche.
What Can Coral Reefs Teach Us?
Coral reefs and coral islands hold secrets which are worthy of note. Firstly, there are time implications involved in view of the supposed slow rate of coral growth. Secondly, they can also tell a story with regard to the events that may have shaped our present oceans and, by implication, the continents as well. Thirdly, fossil reefs can present a challenge to the flood model since their presence is seen to favor slow development over millions of years. Dr. Ariel Roth of the Geoscience Research Institute in California has done extensive research on coral reefs and has published a number of papers which throw light on some of these baffling questions.36, 37
Coral reefs consist of a hard core built up by living organisms that can resist the wave action of the oceans and the reef in general represents one of the most complex marine ecological systems. A reef can trap ocean sediments, and the total structure can thus be quite complex, with part of the reef being constructed by the reef organisms themselves and the rest filled in from other sources.
Fossil reefs in the stratigraphic record thus appear to sug- gest long periods of stability whilst these reefs grew during those time periods in the earth’s history. Fossil reefs can serve as a good trap for oil and they are thus of commercial interest as well. There is, thus, much interest in these reefs and hundreds of fossil reefs are reported throughout the geological column. The question is, how did these fossil reefs form and were they built by reef organisms in the same way as they are being constructed today? If so, then they would seriously challenge the Biblical paradigm of a destruction of the earth by the flood. The fossil reefs could, however, also be allochthonous reefs (formed by transported sedi- ments) and not necessarily be autochthonous reefs (reefs formed by biological processes requiring a long time period). The fossil reefs are generally different to present day reefs since they are often much smaller (thicknesses ranging in the meter range) and the organisms are also different. The interpretation as to whether the corals in the fossil reefs grew there or were transported to their present position catastrophically is similar to that encountered in the study of the fossil trees in the petrified forests. The diversity of organisms (organisms not associated with reefs) and their orientation in the fossil reefs suggests an allochthonous origin, which would be consistent with the catastrophic flood model just as we saw in the case of the petrified forests.
The great Permian reef complex, which is 700 kilometers long and 200 kilometers wide, has long been considered a type of ancient Great Barrier Reef. However, the lack of organisms, which in living reefs construct the frame of the reef, suggests that this reef does not represent a natural reef. Moreover the sedimentary layers associated with the reef suggest a catastrophic
origin. Other fossil reefs such as the Nubrigyn Algal Reefs are now seen as massive debris flows that carried huge blocks of rock as large as one kilometer across and dumped them in their present position. The case for fossil reefs in position of growth is thus the same as for the petrified forests – the reefs are destroyed remnants of pre-flood reefs that were broken up and transported together with other organisms and deposited in layers in the geological column. They are thus recycled reefs and support the catastrophic model. Moreover, they show that the ocean floor was subjected to destructive forces as well and could have played a major role in accounting for some of the features noted in the geological col- umn. The floor of the ocean could have been raised by up-warping during the flood, thus pouring the waters over the continents. The reverse must have happened during the time when the waters were drained off the land to form the present ocean basins. The ques- tion can now be raised – how long did this process take? And this is where living coral reefs and dead coral islands provide some interesting insights.
Living coral reefs are traditionally regarded as slow growers, and they require certain conditions in order to grow. The reefs are produced by a variety of organisms with corals and coralline algae being the main contributors. They are also largely restricted to warmer oceans and, being colonial animals, require symbiotic organisms in order to flourish. Since many of these organisms are algal plants, the reefs must also be in contact with the light and can only grow if the light reaching the reef is suf- ficient to sustain these photosynthetic organisms. Once the coral reef reaches the surface of the water, however, growth is retarded because of the sensitivity of the organisms to ultraviolet radiation and exposure to air. Maximum coral growth is thus achieved a few meters below the surface of the water, but the coral cannot grow from the bottom of the ocean up if the water is too deep (usually 50 meters) to allow for sufficient light to penetrate to that depth. Estimates for the rate of coral growth have usually been derived
from surface studies where growth is not maximal. But high rates have been recorded below the surface of the water, and a number of ships have floundered in charted waters where unexpected reefs had grown rapidly. Growth rates are usually considered to be in the order of 0.8-26 mm/year, but maximum rates of coral growth of up to 414 mm/year have been recorded. Studies also show that if water temperatures are increased by just 5 oC, then growth rates double.
The challenge posed by this information is as follows: How does one account for dead coral islands at the bottom of the oceans in waters that are too deep for coral to grow, and how does one ac- count for tall coral structures that stretch from the ocean floor to the surface. The Great Barrier Reef is 2000 kilometers long and stretches up to 320 kilometers offshore and it is 200 meters thick. This does not pose any serious questions in terms of the time required to pro- duce this reef. Drilling operations at Enewetak Atoll in the Western Pacific, however, have shown this reef material to be 1405 meters thick before reaching a basalt rock base. Applying the general rate of coral growth to this reef, it can seem as if many thousands of years would be required to reach this thickness. Using a growth rate of 414 mm/year, however, could account for this structure in less than 3400 years, which fits in well with a Biblical chronology. If ocean temperatures were higher in the past than in the present, and this seems likely, then the time required to reach this thickness would be even less.
The dead corals in deep ocean waters must have been in con- tact with the light at some stage in order for them to have formed at all. Moreover, the very deep corals, such as the Enewetak reef could not have grown from the bottom up, since they would have been cut off from light at those depths. The only other solution is that the ocean floor was higher in the past and that it then dropped down to its present level. The rate at which the ocean floor dropped down to its present position must have been slow enough in some areas for coral to stay alive and keep growing as in the case of the 1405 meter thick Enewetak coral reef. On the other hand, it must have been fast enough in some areas for the now submerged dead corals at the bottom of the Pacific to have died
when cut off from the light. This places a totally different time frame (in the order of thousands of years) on the great geological upheaval that the earth must have experienced. The scientific world also accepts up and down warping of the ocean floor to explain the obvious conti- nental marine incursions, but their time frame is, of course, reckoned in million of years rather than thousands of years. Corals thus provide a model for what happened during and immediately after the flood. The ocean floor was raised, and the waters poured over the land, and this accounts for the massive marine deposits on the continents. After the flood, the new ocean basin formed by down warping of the ocean floor, which was rapid in some areas, and slow in others. This model is further supported by the fact that all continents show ancient coastlines, which were much further inland than the present ones.
The Post-Flood World
The post-flood world must have been very wet, and it is therefore not surprising to find numerous algal deposits with ferns, reeds, reed fish and dragonfly nests in these deposits. Furthermore, the numerous inland basins and relics of giant lakes imply very wet and hazardous conditions. Today’s Great Salt Lake in Utah, USA is about one sixteenth of its original size, and only a fraction of its original depth. In fact, it is estimated that the lake must have been some 230 meters deep at one stage. Collapsed limestone caves from this time period also contain fossils of bats, which date from the post-flood era. Interestingly, these creatures are identical to present-day forms.
After the flood, recolonization of the continents would have commenced, and this must have taken place quite rapidly. Moreover, the continents must still have provided land bridges for the animals to cross. There is also evidence that the conditions were consider- ably warmer than those of today, as indicated by the numerous temperate-adapted fossil plants and animals found in regions cur- rently climatically unsuitable for their existence. Recolonization of
the new earth would have had to take place in such a way as to be consistent with current concepts on ecological succession. It is to be envisaged that the earth must have been relatively unstable in the immediate post-flood era because of changes in the continents and ocean basins which were still ongoing. Moreover, earthquakes associated with the rise of the continents would have provided further chances for regional catastrophic formations by the rapid drainage of inland basins and later by the breaking up of glacial barriers. There is evidence in the youngest geological layers of succession, and even distribution patterns of animals suggest colonization patterns consistent with a Biblical perspective. For example, genetic studies have shown that species colonization routes in Africa and the Americas occurred in a north-south direc- tion, and in Asia from a west-east direction. (See Figure 5.12)
Once recolonization had largely been completed, ecologi- cal barriers caused by desertification, mountain uplift, or climatic changes could have separated populations. An example here could be the African and Indian elephant populations, which are prob- ably relics of a larger population with its origin midway between the two, a scenario to be expected if the story of Noah’s ark has any credence. Animals would have redistributed themselves quite rapidly, but in view of his great generation length, man would have been the last organism to spread across the new world. Even in spite of man’s long generation time, the time frame for the human population to have grown to present levels is more than adequate to explain current human population levels. In fact, there must have been a number of additional epidemics besides the known ones to explain the low level of the current human population. Growth rates of only 0.5% per year (more than 4 times lower than present rates) would have been sufficient to produce the present human population since the time of the flood some 4500 years ago according to the Scriptures.
Subsequent to the reoccupation of the post-catastrophic world, the earth must have experienced a further cycle of cata- strophism. Evidence for this can be seen in the vast volcanic
deposits present in the youngest layers of the geological column. Volcanism of this scale was probably unleashed when the super- continent “Pangaea” split up to form the present-day continents. It is estimated that 50 000 volcanoes (The mid-Atlantic Ridge and The Pacific Ring of Fire) were active at this time. The volcanoes spewed vast amounts of volcanic ash into the air, thus shielding the sun and causing a rapid drop in temperature. Even relatively minor present-day volcanoes influence global weather patterns after eruption. The combination of warm oceans and sudden drop in radiant energy is ideal for the formation of glaciers, as it can provide the necessary sustainable precipitation to allow glacial advance. This could have induced the Ice Age.
Studies at the Athabasca glacier and glacial bed F26 show that complete glaciation and deglaciation need not have taken more than 600 years. Moreover, evidence for more than one ice age is scant in the geological record, and most of the assumed features in the geological column ascribed to ancient ice ages could equally well have been caused by catastrophic mudflows. The glaciers formed during the Ice Age would have separated populations such as the woolly mammoth into a north- ern and southern population, but conditions must still have been relatively warm in coastal areas, as evidenced by coexistence of animals adapted to different climatic conditions (disharmonious species distribution). An example would be hippopotami and reindeer coexisting in England at that time, as evidenced from the fossil record.
In Siberia, woolly mammoths are found in the ice in such a state of preservation that the flesh is intact. They must therefore have been buried and frozen instantly because such large animals have large heat stores and would have rotted had they not been frozen rapidly. Moreover, they are found in deposits known as Muck. Muck is a geological mystery. It covers one-seventh of the earth’s land surface, is in excess of 4000 feet thick in places and surrounds the Arctic Ocean. It is difficult to explain where it came from, since it occupies flat terrain with no surrounding mountains
from which the muck could have eroded. Oil prospectors have even discovered frozen tree chunks in the muck, and submerged tropical forests up to 1700 feet down have been recorded as well. The muck, now frozen solid, contains the remains of thousands of mammoths and other mammals and shows that they must have succumbed rapidly to a calamity which involved freezing. Judg- ing from plants and animals buried with them, their surroundings must have been more temperate and moist in the past. The animals also found in the muck are rhinoceroses, tigers, bison, horses, antelope, fruit trees, and temperate species of grasses, as well as burrowing animals, such as voles, that could not have burrowed in rock-hard permafrost. It therefore seems logical to propose that the muck originated from flood deposits (particularly since they contain submerged forests) and that the freezing of these animals occurred during the Ice Age events some time after the flood. This timing would allow for population numbers to have reached the levels necessary to account for the numbers associated with the Mammoth remains that have been found or estimated.
Generally, the mammals of the Cenozoic layers are depicted as bizarre and very different to the animals existing today. Common illustrations are those of the sabre-toothed tiger, giant land sloths, and the woolly mammoth. A feature of these animals is their size and diversity. Of all the sabre-toothed tigers removed from the tar pits at Rancho La Brea, the long-toothed variety represents an extreme of the range. Obviously, in a post- catastrophic world with low population densities, the potential for variation would have been great until increase in population density increased the selective pressures and weeded out the ex- tremes of the range. The supposition that these large mammals existed very long ago and must have represented ancient ancestral forms of modern mammals is not consistent with the facts, since pelts of these animals are still found in caves where skeletons of these creatures have been found. In South America, pelts have been found of giant sloths indicating a very recent existence. Today we know that reduction in size need not take millions of
years, but can be achieved rapidly by increases in competition or changes in climate. Animals on islands frequently undergo large- scale reduction in size within the space of a few generations. The potential for rapid change must therefore exist in their gene pool.
Human Evolution
Human evolution is a contentious subject and is governed more by the opinions of the researchers than by the facts reflected in the fossil record. Contentions amongst researchers have led to acrimonious accusations which throw little light on the actual phenomena contended. David Pilbeam, from Yale and Harvard summed the situation up as follows:
I have come to believe that many of the statements we make about the hows and whys of human evolution say as much about us, the paleoanthropologists and the larger society in which we live, as about anything that ‘really’ happened.38
The fossils, upon which human lineages are based, are relatively scarce, and one wonders why so few human fossils have been found. Humans are gregarious, and the bulk of the population would have tended to live close together. Legends concerning pre-flood civilizations and master races abound in folklores around the world, and the legend of the lost continent of Atlantis is one that to this day is generally held in high esteem even in occult circles. These sources believe that Atlantis rep- resents the antediluvian world and that this world disappeared under the sea. Be that as it may, the possibility thus exists that the bulk of human fossils could now be buried in sediments at the bottom of the ocean.
The evidence for man’s evolution is extremely scant, and the main role players are Australopithecines, which are small to medium sized ape-like creatures, which some researchers believe to have walked upright. Remains of these creatures have been found in eastern and southern Africa and the famous Lucy belongs to this group. Then there are the so-called archaic Homo sapiens, which include forms such as the Neanderthals. It is noteworthy that all the forms on which human evolution is based were contemporaneous, which means that they lived at the same time. In fact, all the primates existed at the same time, and the evolutionary tree is once again a morphological sequence pieced together by the scientists according to their perceptions. Australopithecines thus existed side by side with humans and are thus regarded as a side branch from which information can be inferred but that could not have been on the ancestral line of modern humans.
As more information becomes available, more and more intermediary forms are removed from the family tree. Australo- pithecus, as the name suggests, was an ape, but supposedly walked upright. This conclusion is based on footprints in stone that were discovered at Laetoli in Tanzania, which were supposedly made by Lucy-types. However, ‘Lucy-types’ had curled toes and the same knuckle walking wrist anatomy as chimpanzees and goril- las.39 Everything about australopithecines points to the stooped gait of a rolling knuckle-walking chimp-like creature. CAT scans of their inner ear canals (which reflect posture), and their long curved fingers and toes also show that they did not walk upright. The evolutionist Dr. Russel Tuttle of the University of Chicago studied the footprints made by a tribe of people in Peru who walk
barefoot. Based on casts that were made from their footprints, he concluded that the Laetoli prints are in fact identical to those made by humans who habitually walk barefoot. Considering that humans and australopithecines lived at the same time, the most logical conclusion would then seem to be that humans made the prints. Some elements in the literature even suggest that the variation between the three forms (Australopithicus afarensis, A. africanus, A. robustus) is no greater than the normal intra-specific variation experienced in modern ape species.
Lucy is another intermediary under fire, and recent state- ments in the journal New Scientist suggest that she resembled the pygmy chimp. The debate that raged between Johanson and Leaky concerning the suitability of either of these fossils as an intermedi- ary between man and the ape ancestors underlines the uncertainty and scantiness of the information on which bold scientific sup- positions are based. Evidence for Lucy’s intermediary status is extremely fragmentary and cannot stand up to serious scientific scrutiny. Indeed, there is no evidence that these creatures were anything other than apes. The evidence for Lucy’s intermediary status is based on arm to leg length ratios that are supposedly midway between those of apes and man, but in the light of the fragmentary nature of the bones available for study, such a ratio cannot even be determined. The hip is apelike but is claimed to be distorted when there is no ‘undistorted’ hip to compare it with, so on the basis of the evidence it must be an ape hip. The knee of the Lucy fossil, which is also used to substantiate an upright gait, was not even found together with Lucy.
Neanderthal man is also shrouded in controversy, but these fossils show no other evidence than that of Neanderthal being fully human, with a larger brain capacity even than mod- ern man. Homo erectus and Neanderthals are basically the same but the Neanderthal fossils show evidence of bone deformity produced by rickets, which could be the result of conditions that prevailed in the post-flood Ice Age. Of course, the time frames on which all these speculations are based are also not in accord-
ance with the catastrophic model, but then there has been such a huge disparity of dating attempts on the hominid fossils that a discussion on the issue is not even warranted, particularly since the dating techniques are open to question.
Interestingly, there has been an ongoing debate as to the origin of man. Did man originate in Africa or Asia? Protagonists of the two viewpoints have been at loggerheads since the first fossils of so-called human ancestors were discovered on the two continents. In recent times, the theory that man evolved in Africa held sway, but recent evidence again supported the view that man evolved in Asia. The compromise viewpoint that man appeared almost simultaneously in both regions is even more surprising. Again it can be argued that the data is consistent with a distribution from an area midway between the two disputed regions, making the story of Noah’s ark even more plausible.
In his book ‘Bones of Contention: controversies in the search for human origins’,40 Roger Lewin, research news editor of the journal Science at the time, sums up the main issues of contention in the saga of human origins. He sites competition amongst researchers and their passions as some of the reasons for the confusion in the field of hominid evolution. The main con- troversies in his report revolve around: 1) the Taung child found in South Africa and originally rejected but now accepted as an intermediary ancestor in man’s evolution; 2) the Piltdown Hoax where a human skull and an Orangutan’s jaw were ‘doctored’ to lend credence to the naturalistic origin of man and which was unchallenged for almost four decades; 3) Nebraska man who was based on a tooth which turned out to be a pig’s tooth; 4) the distortions towards primitiveness in the original description of Neanderthals; 5) the battle over the dethroning of Ramapithecus from the level of human ancestor to a relative of the orangutan;
6) the heated controversy over the dating of the volcanic layer associated with the hominid fossils in East Africa; 7) the contro- versy between Richard Leakey and Donald Johanson over the
position of the newer australopithecine finds; 8) the conflicts about what constituted the force that brought about human evolutionary change (was it predation, hunting, or cooperation). Lewin’s own feelings are revealed in the following statement:
In the physical realm, any theory of human evolution must explain how it was that an apelike ancestor, equipped with powerful jaws and long, daggerlike canine teeth and able to run at speed on four limbs, became transformed into a slow, bipedal animal whose natural means of defense were at best puny. Add to this the power of intellect, speech, and morality, upon which we ‘stand raised as upon a mountain top’ as Huxley put it, and one has the complete challenge to evolutionary theory.
REFERENCES
1. Herbert Spencer Robinson and Knox Wilson, Myths and Legends of All Nations (New York: Bantam Books, 1950).
2. Derek Victor Ager, The Nature of the Stratigraphic Record, 2nd ed. (London MacMillan Press Ltd., 1983).
3. George Gaylord Simpson, The Meaning of Evolution: A Study of the History of Life and of its Significance for Man (New Haven: Yale University Press, 1949): 326.
4. Tammy Tosk, “Foraminferas in the Fossil Record: Implications for an Ecological Zonation Model,” Origins 15 (1) (1988): 8-18. http://www.grisda.org/origins/15008.pdf
5. Jeffrey S. Levinton, “The Big Bang of Animal Evolution,” Scientific
American (November 1992).
The Genesis Conflict
6. Charles Darwin, The Origin of Species, (New York: Heritage Press, 1963).6. Darwin, Charles (1963) The Origin of Species, Heritage Press (reprint), New York
7. S. Bowring from M.I.T. and Harvard’s Knoll in Time Magazine
(December 1995).
8. Time Magazine (December 4, 1995).
9. D. G. Shu et al., “Lower Cambrian Vertebrates from South China,”
Nature 402 (1999): 42-46.
http://www.efiko.org/material/Lower%20Cambrian%20Verte- brates%20from%20South%20China%20D-G.%20Shu.pdf
10. Stephen Jay Gould, “Return of Hopeful Monsters,” Natural His- tory 86 (June/July): 22-30. http://www.andrew.cmu.edu/user/jksadegh/A%20Good%20Athe- ist%20Secularist%20Skeptical%20Book%20Collection/Gould_ The_Return_of_Hopeful_Monsters_sec.pdf
11. Ann-Sofie Rasmussen and Ulfur Arnason, “Phylogenetic Studies of Complete Mitochondrial DNA Molecules Place Cartilaginous Fishes Within the Tree of Bony Fishes,” Journal of Molecular Evolution 48(1) (1999): 118-123.
12. R. S. Watkins, et al., “Archaeopteryx – A photographic study,” British Journal of Photography (March 18, 1985) and the two sub- sequent issues.
13. Alfred Sherwood Romer, Genetics, Palaeontology and Evolu- tion (Atheneum, 1963).
14. George Gaylord Simpson, The Major Features of Evolution
(Touchstone, 1967).
15. O. C. Marsh, American Journal of Science (1879).
3 – The Fossil Record
16. O. C. Marsh, “Recent Polydactyle Horses,” American Journal of Science 43 (1892): 339-355.
http://marsh.dinodb.com/marsh/Marsh%201892%20-%20Re- cent%20polydactyle%20horses.pdfYork and London; Columbia University Press, p. 263.
17. Dr. Niles Eldrege, curator at the American Museum of Natural History, qtd. in Luther Sunderland, Darwin’s Enigma: Fossils and Other Problems, 4th ed. (Santee: Master Books, 1988): 78. http://www.creationism.org/books/sunderland/DarwinsEnigma/
18. George Gaylord Simpson, The Major Features of Evolution (New York: Columbia University Press, 1963): 263.
19. B. J. Stahl, Vertebrate History: Problems in Evolution (New York, Mc.Graw-Hill, 1974): 489.
20. D. Miller, Seals and Sea Lions (Stillwater, MN: Voyageur Press, 1998): 7.
21. “Hawaiian Monk Seal.” >http://www.kilaueapoint.org/education/ naturefocus/hnf13/index.html>Hawaii Nature Focus, No. 13.
22. John J. Flynn, “Ancestry of sea mammals,” Nature 334(6181) (1988): 383-384.
23. Andre R. Wyss, “Evidence from Flipper Structure for a Single Origin of Pinnipeds,” Nature 334(6181)(1988): 427-428.
24. C.O. Dunbar, Historical Geology (New York: John Wiley and
Sons Inc., 1957).
25. R. H. Dott and R. L. Batten, Evolution of the Earth (New York: Mc.Graw-Hill, 1971): 359.
26. K.O. Stanley, W. M. Jordan, and R. H. Dott, “New Hypothesis of Early Jurassic Paleography and Sediment
The Genesis Conflict
Dispersal for Western United States,” American Association of Petroleum Geologists Bulletin 55 (1971): 10-19.
27. Leonard R. Brand, “Footprints in the Grand Canyon,” Origins
5(2) (1978): 64-82.
http://www.grisda.org/origins/05064.pdf25. Brand, L. 1978. Foot- prints in the Grand Canyon. Origins, 5(2):64-82.
28. R. H. Dott, and R. L. Batten, Evolution of the Earth (New York: Mc.Graw-Hill, 1971): 359.
29. K.O. St anle y, W. M. Jorda n, a nd R. H. Dot t, “New Hypothesis of Early Jurassic Paleography and Sediment Dispersal for Western United States,” American Association of Petroleum Geologists Bulletin 55 (1971): 10-19.
30. Leonard R. Brand, “Footprints in the Grand Canyon,” Origins
5(2) (1978): 64-82. http://www.grisda.org/origins/05064.pdf
31. Leonard R. Brand and James Florence, “Stratigraphic Distribu- tion of Vertebrate Fossil Footprints Compared with Body Fossils,” Origins, 9(2): 67-74. http://www.grisda.org/origins/09067.pdf
32. Ibid.
33. D. M. Martill, D. J. Batten and D. K. Loydell, “A New Specimen of the Thyreophram Dinosaur cf. Scelidosaurus with Soft Tissue Preservation,” Palaeontology 43 (2000): 549-559.
34. Luis M. Chiappe et al., “Sauropod Dinosaur Embryos from the Late Cretaceous of Patagonia,” Nature 396(6708) (1998): 258-61.
35. >http://www.canoe.com/CNEWSScienceArchive/981117_dino- saurs.html> May 25, 2000.
36. A. C. Burke and A. Feduccia, “Developmental Patterns and the
- – The Fossil Record
Identification of Homologies in the Avian Hand,” Science 278 (5338) (1997): 666-8, with a perspective by R. Hinchliffe, “The Forward March of the Bird-Dinosaurs Halted?” (596-97).
37. Quoted in Ann Gibbons, “Lung fossils suggest dinos breathed in cold blood,” Science 278(5341) (1997): 1229-1230. Ruben’s paper was published in the same issue, “Lung ventilation in Theropod Dinosaurs and Early Birds,” 1267-1270.
38. Harold G. Coffin, “Orientation of Trees in Yellowstone Petrified
Forests,” Journal of Paleontology 50(3) (1976) :539-543.
39. Time Magazine (December 4, 1995).
40. Harold G. Coffin, “Erect Floating Stumps in Spirit Lake, Wash- ington,” Geology 11 (1983): 298-299.
41. Time Magazine (December 4, 1995).
42 P. McCafferty, “Instant Petrified Wood?” Popular Science (October 1992): 56-57.
43. Roy Piggott, The Australian Lapidary Magazine (January 1970): 9.
44. Ariel A. Roth, “Fossil Reefs and Time,” Origins 22(2) (1995): 86-104.
http://www.grisda.org/origins/22086.pdf
45. Ariel A. Roth, “Coral Reef Growth,” Origins 6(2) (1995): 88-95. http://www.grisda.org/origins/06088.pdf
46. D. Pilbeam, “Rethinking Human Origins,” Discovery 13(1): 2-10.
47. Mark Collard and Leslie C. Aiello, “Human Evolution: From Forelimbs to Two Legs,” Nature 404(6776) (2000): 339-340 and Brian G. Richmond and David S. Strait, “Evidence that Humans
The Genesis Conflict
Evolved from a Knuckle-Walking Ancestor,” Nature 404(6776) (2000): 382-385.
48. Roger Lewin, Bones of Contention: Controversies in the Search for Human Origins (New York: Simon and Schuster, 1987): 312- 313.
I have read so many posts about the blogger lovers however this post is really a good piece of writing, keep it up
I have read so many posts about the blogger lovers howeverthis post is really a good piece of writing, keep it up.
I have read so many posts about the blogger lovers however this post is really a good piece of writing, keep it up
Here at this site really the fastidious material collection so that everybody can enjoy a lot.
We are tied directly into the sate’s renewal database which allows us to process your request almost instantly
wow… what a great blog, this writter who wrote this article it’s realy a great blogger, this article so inspiring me to be a better person
https://www.philadelphia.edu.jo/library/directors-message-library